Artbildung
Bei der Speziation geht es darum, wie sich Arten bilden. Sie ist ein wichtiger Teil der Evolutionsbiologie. Darwin ging davon aus, dass die meisten Arten direkt aus bereits existierenden Arten entstanden sind. Dies wird Anagenese genannt: Arten…
Bei der Speziation geht es darum, wie sich Arten bilden. Sie ist ein wichtiger Teil der Evolutionsbiologie.
Darwin ging davon aus, dass die meisten Arten direkt aus bereits existierenden Arten entstanden sind. Dies wird Anagenese genannt: Arten durch Veränderung oder "phyletische Evolution". Während eines Großteils des 20. Jahrhunderts dachten wir, die meisten Arten seien durch vorherige Artenaufspaltung entstanden: Cladogenese. Die allgemeine Ansicht war, dass die meisten Artenspaltungen durch isolierende Mechanismen verursacht oder auf ihrem Weg unterstützt werden.
Zweifellos ist die physische Trennung von Arten, die einst zusammenlebten, ein Hauptfaktor. Er wird durch so viele Beispiele veranschaulicht, von denen einige im Folgenden erörtert werden.
Die Arbeit der letzten 20 Jahre hat jedoch einige andere Ursachen aufgezeigt. Die Analyse der DNA-Sequenz von Lebewesen hat gezeigt, dass es häufig zu einer gewissen Hybridisierung zwischen verwandten Arten kommt. Das bedeutet, dass Gene durch diese Kreuzungen übertragen worden sind. Das wiederum bedeutet, dass die reproduktive Isolation nicht die einzige Definition einer Spezies ist, und die Speziation erfordert nicht immer Allopatrie (Spezies, die reproduktiv getrennt werden müssen). Die folgenden Abschnitte veranschaulichen die Idee, dass die physische Trennung bei der Bildung neuer Arten von größter Bedeutung war.
Bildergalerie
10 Bilder









Isolierende Mechanismen
Isolierende Mechanismen sind Dinge, die eine erfolgreiche Fortpflanzung zwischen Gruppen innerhalb einer Art verhindern. Die reproduktive Isolation von Populationen ist etabliert. Dies ist für das biologische Artkonzept besonders wichtig, da Arten durch reproduktive Isolation definiert werden.
Isolierende Mechanismen können in zwei Gruppen eingeteilt werden, vor und nach der Befruchtung.
Vor der Befruchtung
Faktoren, die Individuen von der Paarung abhalten.
- Geografische Isolation: Die Arten kommen in verschiedenen Gebieten vor und sind oft durch Barrieren getrennt.
- Zeitliche Isolation: Individuen paaren sich nicht, weil sie zu unterschiedlichen Zeiten aktiv sind. Dies können verschiedene Tageszeiten oder verschiedene Jahreszeiten sein. Die Paarungszeiten der Arten stimmen möglicherweise nicht überein. Die Individuen begegnen einander nicht, weder während der Paarungszeit noch überhaupt nicht.
- Ökologische Isolation: Individuen paaren sich nur in ihrem bevorzugten Lebensraum. Sie treffen nicht auf Individuen anderer Arten mit anderen ökologischen Präferenzen.
- Verhaltensbedingte Isolation: Individuen verschiedener Spezies können aufeinander treffen, aber man erkennt keine sexuellen Hinweise, die gegeben werden könnten. In den meisten Fällen wählt ein Individuum ein Mitglied seiner eigenen Spezies aus.
- Mechanische Isolierung: Es kann eine Kopulation versucht werden, aber es findet keine Übertragung von Spermien statt. Die Individuen können aufgrund ihrer Größe oder Morphologie inkompatibel sein.
- Gametische Inkompatibilität: Es findet ein Spermientransfer statt, aber die Eizelle wird nicht befruchtet.
Nach der Befruchtung
Faktoren, die eine erfolgreiche Paarung verhindern, wie genetische Unverträglichkeit, Hybriduntauglichkeit oder Sterilität.
- Zygotische Mortalität: Die Eizelle wird befruchtet, aber die Zygote entwickelt sich nicht.
- Hybride Lebensfähigkeit: Ein hybrider Embryo bildet sich, ist aber nicht lebensfähig.
- Hybridsterilität: Die Hybride ist lebensfähig, aber der resultierende Erwachsene ist steril.
- Hybrider Zusammenbruch: Hybriden der ersten Generation (F1) sind lebensfähig und fruchtbar, aber weitere Hybridgenerationen (F2 und Rückkreuzungen) sind nicht lebensfähig oder steril.

Geografische Isolation
Man geht davon aus, dass dies die häufigste Ursache für Speziation ist. Der erste Mensch, der daran dachte, war Moritz Wagner, ein deutscher Forscher und Naturhistoriker.
Wagners frühe Karriere war die eines Geographen, und er veröffentlichte eine Reihe von geographischen Büchern über Nordafrika, den Nahen Osten und das tropische Amerika. Er war auch ein begeisterter Naturforscher und Sammler, und für diese Arbeit ist er unter Biologen am besten bekannt. Ernst Mayr, der Evolutionist und Historiker der Biologie, hat über die Bedeutung Wagners berichtet. S. 562-565
Während seines dreijährigen Algerienaufenthaltes untersuchte Wagner (neben anderen Aktivitäten) die flugunfähigen Käfer Pimelia und Melasoma. Jede Gattung ist in eine Reihe von Arten unterteilt, von denen jede auf einen Abschnitt der Nordküste zwischen Flüssen, die vom Atlasgebirge zum Mittelmeer hinabfließen, beschränkt ist. Sobald man einen Fluss überquert, erscheint eine andere, aber eng verwandte Art.
"... eine beginnende Spezies wird nur [entstehen], wenn einige wenige Individuen die Grenzen ihres Verbreitungsgebietes überschreiten... die Bildung einer neuen Rasse wird niemals gelingen... ohne eine lange fortgesetzte Trennung der Kolonisten von den anderen Mitgliedern ihrer Spezies".
Dies war eine frühe Beschreibung eines Prozesses einer Art von geographischer Speziation. Im Jahre 1942 wurde sie von Mayr wieder eingeführt, und die Bedeutung der geographischen Speziation wurde zu einem der Kerngedanken der evolutionären Synthese.
Ein anderer Begriff für geographische Speziation ist allopatrische Speziation. Allopatrie bedeutet 'anderes Land'.
Leere Inseln
Vulkanische Inseln entstehen ohne Leben, und alles Leben muss durch Wind oder Wasser getragen ankommen. Von den Hawaii-Inseln und den Galapagos-Inseln wissen wir, dass sich alle Lebensformen verändern, wenn sie vom Festland auf die Inseln gelangen.
Auf etwa 6.500 Quadratmeilen (17.000 km2) haben die Hawaii-Inseln die artenreichste Sammlung von Drosophilidfliegen der Welt, die von Regenwäldern bis zu Bergwiesen leben. Etwa 800 hawaiianische Drosophilidenarten sind bekannt.
Studien zeigen einen deutlichen "Fluss" von Arten von älteren zu neueren Inseln. Es gibt auch Fälle von Kolonisierung zurück auf ältere Inseln und Überspringen von Inseln, aber diese sind viel seltener.
Nach der radioaktiven Datierung mit Kalium/Argon datieren die heutigen Inseln von vor 0,4 Millionen Jahren (mya) (Mauna Kea) bis 10mya (Necker). Das älteste noch über dem Meer liegende Mitglied des hawaiianischen Archipels ist das Kure-Atoll, das auf 30 mya datiert werden kann.
Der Archipel selbst, der durch die Bewegung der pazifischen Platte über einen Hot Spot entsteht, existiert schon viel länger, zumindest bis in die Kreidezeit hinein. Die hawaiianischen Inseln und die ehemaligen Inseln, die jetzt unter dem Meer liegen, bilden die Seamount-Kette Hawaiian-Emperor; und viele der Unterwasserberge sind Guyots.
Alle einheimischen Drosophilid-Arten auf Hawaiʻi stammen offenbar von einer einzigen Vorfahrenart ab, die vor etwa 20 Millionen Jahren die Inseln kolonisiert hat. Die anschließende adaptive Radiation wurde durch mangelnde Konkurrenz und eine Vielzahl von freien Nischen vorangetrieben. Obwohl es für ein einzelnes schwangeres Weibchen möglich wäre, eine Insel zu besiedeln, ist es wahrscheinlicher, dass es sich um eine Gruppe der gleichen Spezies handelt.
Es gibt andere Tiere und Pflanzen auf dem hawaiianischen Archipel, die ähnliche, wenn auch weniger spektakuläre, anpassungsfähige Strahlungen erfahren haben.
Ring-Arten
In der Biologie ist eine Ringart eine zusammenhängende Reihe benachbarter Populationen, von denen sich jede mit den Nachbarpopulationen kreuzen kann. Die beiden Enden der Kette überlappen sich.
Die beiden Endpopulationen in der Serie sind zu weit von der Kreuzung entfernt. Solche nicht durch Kreuzung, aber genetisch miteinander verbundenen "Endpopulationen" können in derselben Region koexistieren und so einen "Ring" schließen.
Ringarten liefern wichtige Beweise für die Evolution: Sie veranschaulichen, was im Laufe der Zeit geschieht, wenn Populationen genetisch divergieren. Richard Dawkins bemerkte, dass Ringarten "uns in der räumlichen Dimension nur etwas zeigen, was in der zeitlichen Dimension immer geschehen muss".
Es ist jedoch schwierig, ein einfaches, unkompliziertes Beispiel zu finden.
Larus-Möwen
Ein klassisches Beispiel für Ringarten ist der zirkumpolare Artenring der Larus-Möwen. Das Verbreitungsgebiet dieser Möwen bildet einen Ring um den Nordpol, der normalerweise nicht von einzelnen Möwen überflogen wird.
Im Großen und Ganzen können sich die Möwen bis zu einem gewissen Grad mit ihren Nachbarn kreuzen, außer an den beiden Enden des Rings.
Die kleineren Schwarzbandmöwen und Heringsmöwen sind so unterschiedlich, dass sie sich normalerweise nicht miteinander kreuzen; daher bildet die Gruppe der Möwen ein Kontinuum, außer dort, wo sich die beiden Abstammungslinien in Europa treffen.
Obwohl die Realität viel komplizierter ist, zeigt sie die allgemeine Idee gut.
Ensatina-Salamander
Der Ensatina-Salamander ist eine Ringart in den Bergen um das kalifornische Central Valley. Der Komplex bildet eine Hufeisenform um die Berge. Obwohl es zwischen jeder der 19 Populationen um das Hufeisen herum zu Kreuzungen kommen kann, kann sich die Unterart Ensatina eschscholtzii am westlichen Ende des Hufeisens nicht mit der Unterart Ensatina klauberi am östlichen Ende des Hufeisens kreuzen. Es ist eine Illustration "fast aller Stadien eines Speziationsprozesses" (Dobzhansky). Richard Highton argumentierte, dass Ensatina ein Fall von mehreren Arten ist und nicht ein Kontinuum einer Art.
Der Grünliche Grasmücke
Der Grüne Grasmücke (Phylloscopus trochiloides) hat eine Reihe von Unterarten, von denen P. t . viridianus die in Europa bekannteste ist. Es handelt sich um eine Ringart mit Populationen, die östlich und westlich der tibetischen Hochebene auseinandergehen und sich später auf der nördlichen Seite treffen. Ihre Verwandtschaft ist ziemlich verwirrend.




Sympatrische Speziation
Sympatrische Speziation bezieht sich auf die Bildung von zwei oder mehr nachkommenden Arten von einer einzigen angestammten Art, die alle den gleichen geographischen Standort besetzen. Man geht heute davon aus, dass dies sehr häufig vorkommt.
Bei der sympatrischen Speziation divergieren die Arten, während sie den gleichen Ort bewohnen. Häufig zitierte Beispiele für sympatrische Speziation finden sich bei Insekten, die von verschiedenen Wirtspflanzen im selben Gebiet abhängig werden.
Die Existenz der sympatrischen Speziation als Mechanismus der Speziation wurde heftig umstritten. Es wurde argumentiert, dass die Beweise für sympatrische Speziation in der Tat Beispiele für mikrogeografische Speziation seien. Im Allgemeinen ist dies jetzt nicht die bevorzugte Erklärung. Vielmehr ist sie das Ergebnis einer Hybridisierung zwischen eng verwandten Arten, gefolgt von einer natürlichen Selektion, die auf die Nachkommen solcher Kreuzungen hinarbeitet. Ein weithin akzeptiertes Beispiel für eine sympatrische Speziation sind die Buntbarsche des Nabugabo-Sees in Ostafrika, von denen man annimmt, dass sie durch sexuelle Selektion entstanden sind.
Speziation über Polyploidisierung
Die Polyploidie hat viele schnelle Speziationsereignisse verursacht, weil Nachkommen von z.B. tetraploiden x diploiden Paarungen oft triploide sterile Nachkommen hervorbringen.
Allerdings werden nicht alle Polyploide reproduktiv von ihren Elternpflanzen isoliert, und der Genfluss kann immer noch stattfinden, zum Beispiel durch triploide Hybrid x diploide Paarungen, die Tetraploide produzieren.
Viele der existierenden Pflanzen- und die meisten Tierarten haben in ihrer Evolutionsgeschichte offensichtlich eine Polyploidisierung durchgemacht. Die Fortpflanzung erfolgreicher polyploider Arten ist manchmal ungeschlechtlich, durch Parthenogenese. Aus unbekannten Gründen sind viele ungeschlechtliche Organismen polyploid.
Weißdornfliege
Ein Beispiel für die Evolution bei der Arbeit ist der Fall der Weißdornfliege, Rhagoletis pomonella, die sich anscheinend einer sympatrischen Speziation unterzieht.
Verschiedene Populationen der Weißdornfliege ernähren sich von verschiedenen Früchten. Eine ausgeprägte Population entstand in Nordamerika im 19. Jahrhundert, einige Zeit nachdem Äpfel, eine nicht einheimische Art, eingeführt worden waren. Diese sich von Äpfeln ernährende Population ernährt sich normalerweise nur von Äpfeln und nicht von der historisch bevorzugten Frucht des Weißdorns. Die heutige Weißdorn-Futterpopulation ernährt sich normalerweise nicht von Äpfeln.
Einige Hinweise deuten darauf hin, dass eine sympatrische Speziation stattfindet. Weißdornfliegen werden später in der Saison reif und brauchen länger zur Reife als Apfelfliegen; und es gibt kaum Anzeichen für eine Kreuzung (Forscher haben eine Hybridisierungsrate von 4-6% dokumentiert).
Das Aufkommen der neuen Weißdornfliege ist ein Beispiel für die fortschreitende Evolution.
Hybridisierung
Selten bildet sich eine neue Art, wenn sich einzelne Mitglieder verschiedener Arten paaren. In der Regel sind die Produkte solcher Hybridpaarungen unfruchtbar (nicht fruchtbar) oder relativ unfruchtbar und werden daher durch natürliche Selektion eliminiert.
Es wurde ein Beispiel für eine neue und erfolgreiche Hybridart gefunden. Bei der neuen Art handelt es sich um einen Hybriden aus dem italienischen und spanischen Spatzen, und die grundlegenden Fakten wurden durch Sequenzanalyse der DNA in ihrem Blut überprüft. Der Vogel lebt in Italien in einer Region, in der beide Elternarten leben. Er pflanzt sich nicht mit dem spanischen Spatz fort, obwohl er Seite an Seite mit ihnen lebt.
Künstliche Speziation
Neue Arten sind durch domestizierte Tierhaltung entstanden, aber die ersten Daten und Methoden sind nicht klar. Zum Beispiel sind Hausschafe durch Hybridisierung entstanden und produzieren keine lebensfähigen Nachkommen mehr mit Ovis orientalis, einer Art, von der sie abstammen.
Domestizierte Rinder hingegen können als dieselbe Art wie mehrere Arten von Wildochsen, Gaur, Yak usw. angesehen werden, da sie mit ihnen leicht fruchtbaren Nachwuchs produzieren.
Labor-Arten
Die am besten dokumentierten Kreationen neuer Arten im Labor wurden in den späten 1980er Jahren durchgeführt. William Rice und G.W. Salt züchteten Fruchtfliegen, Drosophila melanogaster, unter Verwendung eines Labyrinths mit drei verschiedenen Lebensräumen, wie hell/dunkel und nass/trocken.
Jede Generation wurde in das Labyrinth gesetzt, und die Fliegengruppen, die aus zwei der acht Ausgänge herauskamen, wurden getrennt, um in ihren jeweiligen Gruppen miteinander zu züchten. Nach fünfunddreißig Generationen wurden die beiden Gruppen und ihre Nachkommen aufgrund ihrer starken Habitatpräferenzen reproduktiv isoliert: Sie paarten sich nur innerhalb der von ihnen bevorzugten Gebiete und paarten sich daher nicht mit Fliegen, die die anderen Gebiete bevorzugten. Die Geschichte solcher Versuche wird in Rice und Hostert (1993) beschrieben.
Diane Dodd konnte auch zeigen, wie sich bei Drosophila pseudoobscura nach nur acht Generationen unter Verwendung verschiedener Futterarten, Stärke und Maltose, eine reproduktive Isolation aus Paarungspräferenzen entwickeln kann.
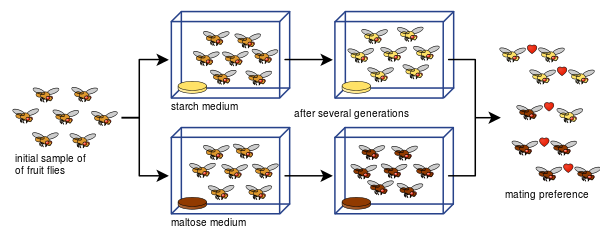
Dodds Experiment war für viele andere leicht zu wiederholen, auch mit anderen Arten von Fruchtfliegen und Lebensmitteln.
Verstärkung (Wallace-Effekt)
Verstärkung ist der Prozess, durch den die natürliche Auslese die reproduktive Isolation erhöht.
Sie kann auftreten, nachdem zwei Populationen derselben Art getrennt wurden und dann wieder in Kontakt gekommen sind. Wenn ihre reproduktive Isolation vollständig war, dann haben sie sich bereits zu zwei getrennten, inkompatiblen Arten entwickelt.
Wenn ihre reproduktive Isolation unvollständig ist, dann werden aus weiteren Paarungen zwischen den Populationen Hybriden entstehen, die fruchtbar sein können oder auch nicht. Wenn die Hybriden unfruchtbar oder fruchtbar, aber weniger fit als ihre Vorfahren sind, dann gibt es keine weitere reproduktive Isolation, und die Speziation ist im Wesentlichen erfolgt (z.B. wie bei Pferden und Eseln).
Dahinter steht die Überlegung, dass, wenn die Eltern der Hybridnachkommen von Natur aus jeweils Merkmale für ihre eigene, unterschiedliche Umgebung ausgewählt haben, die Hybridnachkommen Züge von beiden tragen werden und weder zu einer Nische noch zu einem Elternteil passen würden. Die geringe Fitness der Hybriden würde dazu führen, dass die Selektion assortative Paarungen bevorzugt, was die Hybridisierung reduzieren würde.
Dies wird manchmal als Wallace-Effekt bezeichnet, nach dem Evolutionsbiologen Alfred Russel Wallace, der im späten 19. Jahrhundert vermutete, dass er ein wichtiger Faktor bei der Speziation sein könnte.
Wenn die hybriden Nachkommen fitter sind als ihre Vorfahren, dann verschmelzen die Populationen innerhalb des Gebietes, mit dem sie in Kontakt stehen, wieder zu derselben Art.
Eine Verstärkung ist für die Trennung erforderlich, wenn es eine "Hybridzone" zwischen zwei Formen einer Art gibt. Hybridzonen sind Regionen, in denen divergierende Populationen aufeinander treffen und sich vermischen. Hybride Nachkommen sind in diesen Regionen sehr häufig, die in der Regel dadurch entstehen, dass divergierende Arten in sekundären Kontakt kommen.
Ohne Verstärkung würden sich die beiden Populationen oder Arten unkontrollierbar kreuzen. Verstärkung kann in künstlichen Selektionsexperimenten, wie oben beschrieben, induziert werden.
Fragen und Antworten
F: Was ist Speziation?
A: Speziation ist der Prozess, wie sich Arten bilden. Er ist ein wichtiger Teil der Evolutionsbiologie.
F: Was hat Darwin über die Entstehung von Arten gedacht?
A: Darwin war der Meinung, dass die meisten Arten direkt aus bereits existierenden Arten hervorgegangen sind, was er Anagenese oder 'phyletische Evolution' nannte.
F: Wie wurde die Speziation im 20. Jahrhundert allgemein gesehen?
A: Im 20. Jahrhundert glaubten die Wissenschaftler, dass die meisten Arten durch die Aufspaltung früherer Arten entstanden sind, was als Kladogenese bezeichnet wurde. Man glaubte auch, dass diese Aufspaltung durch isolierende Mechanismen wie die physische Trennung verursacht oder unterstützt wurde.
F: Wie hat die jüngste Forschung unser Verständnis von Artbildung verändert?
A: Neuere Forschungen haben gezeigt, dass es zur Hybridisierung zwischen verwandten Arten kommen kann und Gene durch diese Kreuzungen übertragen werden können. Das bedeutet, dass eine reproduktive Isolation nicht immer notwendig ist, um eine Art zu definieren, und dass Allopatrie (physische Trennung) nicht unbedingt erforderlich ist, um eine Art zu bilden.
F: Was hat die physische Trennung mit der Entstehung neuer Arten zu tun?
A: Man geht davon aus, dass die physische Trennung zwischen ehemals koexistierenden Arten ein wichtiger Faktor bei der Entstehung neuer Arten ist, wie zahlreiche Beispiele in der wissenschaftlichen Literatur zeigen.
Verwandte Artikel
Autor
AlegsaOnline.com Artbildung Leandro Alegsa
URL: https://de.alegsaonline.com/art/92570
Quellen
- doi.org : 10.1126/science.23.587.506
- pubmed.ncbi.nlm.nih.gov : 17789700
- doi.org : 10.1086/279001
- science.siu.edu : "The concept of species"
- abacus.gene.ucl.ac.uk : "Isolating mechanisms"
- mallet.oeb.harvard.edu : mallet.oeb.harvard.edu/publications/what-speciation
- books.google.com : "The Continental Drift Controversy: Introduction of Seafloor Spreading," p. 292
- pubs.usgs.gov : Volcanism in Hawaii, U.S. Geological Survey Professional Paper 1350, pp. 5-54
- rspb.royalsocietypublishing.org : "The herring gull complex is not a ring species"
- doi.org : 10.1098/rspb.2004.2679
- journals.royalsoc.ac.uk : Electronic Appendix
- pnas.org : Incipient species formation in salamanders of the Ensatina complex
- jstor.org : "Is Ensatina eschscholtzii a ring-species?"
- doi.org : 10.1146/annurev.genet.34.1.401
- pubmed.ncbi.nlm.nih.gov : 11092833